Категория: Инструкции
НЕЙРОТЕНЗИН. нейропептид . обладающий гормональным действием. Первичная структура pGlu—Leu—Туr— —Glu—Asn—Lys—Pro—Arg—Arg—Pro—Tyr—He—Leu (мол. м. 1671; букв. обозначения см. в ст. Аминокислоты . р Glu-остаток пироглутаминовой кислоты).
нейротензин присутствует в пикомолярных кол-вах в гипоталамусе, в слизистой тонкого кишечника и в желудке млекопитающих и человека. По своим физиол. свойствам близок к кининам . Обладает сильным гипотензивным действием, вызывает сокращение гладкой мускулатуры, понижает температуру тела, повышает содержание в крови глюкозы и глюкагона. обладает способностью связываться с рецепторами тучных клеток. При действии на гипофиз нейротензин стимулирует секрецию лютеинизирующего гормона и фолликулостимулирующего гормона .
нейротензин получают путем синтеза; впервые его выделили из бычьих гипоталамусов Р. Карравей и С. Лимен в 1973; ими же в 1975 установлена его первичная структура.
Лит.: "Биоорганическая химия", 1980, т. 6, № 3, с. 379-85; Carraway R. Leeman S. "J. Biol. Chem.", 1973, v. 248, № 19, p. 6854-61; Yajima H. [a.o.], "Chem. Pharm. Bull.", 1976, v. 24, № 11, p. 2692-98; Brown D. Miller R.Y. "Ann Rep. Med. Chem.", 1982, v. 17, p. 271-80. B.B. Баев.
Подписка на новости
Новости компанийПодписка на новости

нейропептид, обладающий гормональным действием. Первичная структура pGluЧLeuЧТуrЧ ЧGluЧAsnЧLysЧProЧArgЧArgЧProЧTyrЧHeЧLeu (мол. м. 1671; букв. обозначения см. в ст. Аминокислоты, р Glu-остаток пироглутаминовой к-ты).
Н. присутствует в пикомолярных кол-вах в гипоталамусе, в слизистой тонкого кишечника и в желудке млекопитающих и человека. По своим физиол. св-вам близок к кининам. Обладает сильным гипотензивным действием, вызывает сокращение гладкой мускулатуры, понижает т-ру тела, повышает содержание в крови глюкозы и глюкагона, обладает способностью связываться с рецепторами тучных клеток. При действии на гипофиз Н. стимулирует секрецию лютеинизирующего гормона и фолликулостимулирующего гормона.
Н. получают путем синтеза; впервые его выделили из бычьих гипоталамусов Р. Карравей и С. Лимен в 1973; ими же в 1975 установлена его первичная структура.
Лит.: "Биоорганическая химия ", 1980, т. 6, № 3, с. 379-85; Carraway R. Leeman S. "J. Biol. Chem.", 1973, v. 248, № 19, p. 6854-61; Yajima H. [a.o.], "Chem. Pharm. Bull.", 1976, v. 24, № 11, p. 2692-98; Brown D. Miller R.Y. "Ann Rep. Med. Chem.", 1982, v. 17, p. 271-80. B.B. Баев.
Химическая энциклопедия. — М. Советская энциклопедия. Под ред. И. Л. Кнунянца. 1988 .
Смотреть что такое "НЕЙРОТЕНЗИН" в других словарях:Регуля?торные пепти?ды — высокомолекулярные соединения, представляющие собой цепочку аминокислотных остатков, соединенных пептидной связью. Р. п. насчитывающие не более 20 аминокислотных остатков, называют олигопептидами, от 20 до 100 полипептидами, свыше 100 белками.… … Медицинская энциклопедия
Эндокринная система — Главные железы внутренней секреции (слева мужчина, справа женщина): 1. Эпифиз (относят к диффузной эндокринной системе) 2. Гипофиз 3. Щитовидная железа 4 … Википедия
Апу?д-систе?ма — APUD система (APUD аббревиатура, образованная из первых букв англ. слов amines амины, precursor предшественник, uptake усвоение, поглощение, decarboxylation декарбоксилирование; синоним диффузная нейроэндокринная система) система клеток,… … Медицинская энциклопедия
Гастроинтестина?льные гормо?ны — (греч. gaster желудок + лат. intestinum кишка) группа биологически активных пептидов, вырабатываемых эндокринными клетками и нейронами желудочно кишечного тракта и поджелудочной железы; обладают регуляторным влиянием на секреторные функции,… … Медицинская энциклопедия
РЕГУЛЯТОРНЫЕ ПЕПТИДЫ — (нейропептиды), биологически активные вещества, состоящие из различного числа аминокислотных остатков (от двух до нескольких десятков). Различают олигопептиды, состоящие из небольшого числа аминокислотных остатков, и более крупные полипептиды,… … Энциклопедический словарь
Боль — I тягостное ощущение, отражающее психофизиологическое состояние человека, которое возникает под влиянием сверхсильных или разрушительных раздражителей. Биологическое и физиологическое значение боли состоит в том, что она сигнализирует о наличии… … Медицинская энциклопедия
Кишечник — I Кишечник (intestina) часть пищеварительного канала, начинающаяся от привратника желудка и заканчивающаяся заднепроходным отверстием. В кишечнике происходит переваривание и всасывание пищи, удаление образующихся шлаков, синтезируются некоторые… … Медицинская энциклопедия
ГАСТРОИНТЕСТИНАЛЬНЫЕ ГОРМОНЫ — (от греч. gaster желудок и лат. intestinum кишка), группа биологически активных веществ пептидной природы, вырабатываемых в слизистой оболочке желудочно кишечного тракта у позвоночных; участвуют в регуляции деятельности пищеварит. системы, а… … Биологический энциклопедический словарь
Нейропептиды — Нейропептиды пептиды (разновидность молекул белка), образующиеся в центральной или периферической нервной системе и регулирующие физиологические функции организма человека и животных. Содержание 1 Структура 2 Синтез 3 … Википедия
НЕЙРОТЕНЗИН. нейропептид, обладающий гормональным действием. Первичная структура pGlu—Leu—Туr— —Glu—Asn—Lys—Pro—Arg—Arg—Pro—Tyr—He—Leu (мол. м. 1671; букв. обозначения см. в ст. Аминокислоты, р Glu-остаток пироглутаминовой к-ты).
Н. присутствует в пикомолярных кол-вах в гипоталамусе, в слизистой тонкого кишечника и в желудке млекопитающих и человека. По своим физиол. св-вам близок к кининам. Обладает сильным гипотензивным действием, вызывает сокращение гладкой мускулатуры, понижает т-ру тела, повышает содержание в крови глюкозы и глюкагона, обладает способностью связываться с рецепторами тучных клеток. При действии на гипофиз Н. стимулирует секрецию лютеинизирующего гормона и фолликулостимулирующего гормона.
Н. получают путем синтеза; впервые его выделили из бычьих гипоталамусов Р. Карравей и С. Лимен в 1973; ими же в 1975 установлена его первичная структура.
Лит.: "Биоорганическая химия", 1980, т. 6, № 3, с. 379-85; Carraway R. Leeman S. "J. Biol. Chem.", 1973, v. 248, № 19, p. 6854-61; Yajima H. [a.o.], "Chem. Pharm. Bull.", 1976, v. 24, № 11, p. 2692-98; Brown D. Miller R.Y. "Ann Rep. Med. Chem.", 1982, v. 17, p. 271-80. B.B. Баев.
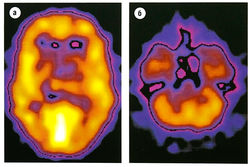

Рис. 8.25 Кортикальное распределение ГАМКд-рецепторных комплексов. Изображение получено с помощью радиоактивно меченного аналога бензодиазепина ломазенила и однофотонной эмиссионной компьютерной томографии. Самые светлые области имеют наивысшую плотность рецепторов, (а) Изображение на уровне среднезатылочной коры, (б) Изображение на уровне мозжечка.
ГАМК — главный тормозной нейромедиатор в ЦНС
ГАМК — аминокислота, действующая прежде всего на ГАМКд- и ГАМКв-рецепторы. ГАМКА-рецепторы присутствуют на 40% нейронов. Кортикальное распределение ГАМКд изображено на рис. 8.25. С1'-канал зависит от ГАМКА-рецептора, в то время как ГАМКв-рецепторы соединены с G-белками.
С нарушениями ГАМК-системы, как полагают,могут быть связаны невротические расстройства. По результатам недавних исследований было высказано предположение о роли ГАМК в этиологии шизофрении.
Глицин — необходимый нейромедиатор для действия глутамата
Глицин необходим для реализации эффектов глутамата. Кроме того, глицин действует на собственные рецепторы, связанные с С1"-каналом и ингибирующие нервные функции.
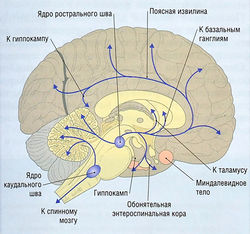
Рис. 8.26 5-Гидрокситриптаминовые пути. 5-НТ-содержащие нейроны найдены в ядрах медиального и дорсального шва, хвостатом и голубоватом пятнах, межножковой области.
АХ действует как нейромедиатор в ЦНС так же, как и на периферии.
Центральное первичное АХ-содержащее ядро -базальное ядро, которое расположено в переднем мозге и идет к коре головного мозга и лимбической системе. Холинергические волокна в ретикулярной системе поступают к коре головного мозга, лимбической системе, гипоталамусу и таламусу.
Идентифицировано девять различных 5-НТ-рецепторов
Наиболее изучены подтипы 5-НТ1А, 5-НТ2В, 5-НТ2с и 5-НТ3. Серотонинергические клеточные тельца в основном локализуются в области верхнего варолиевого моста и среднего мозга. Классические области для 5-НТ-содержащих нейронов — ядра медиального и дорсального шва. Нейроны от ядер шва идут к базальным ганглиям и различным частям лимбической системы и имеют широкое распределение по всей коре головного мозга в дополнение к связям с мозжечком (рис. 8.26).
Все 5-НТ-рецепторы идентифицированы как рецепторы, соединенные с G-белком, кроме подтипа 5-НТ3, который расположен на рецептор-опосредуемом №+/К+-канале.
5-НТ синтезируется из триптофана при участии триптофангидроксилазы, и поступление триптофана лимитирует его синтез. 5-НТ прежде всего метаболизируется моноаминоксидазой А в 5-гидроксииндолацетиловую кислоту (5-ГИАК).
Норэпинефрин широко распространен в ЦНС
Норэпинефрин действует как нейромедиатор в ЦНС и ВНС (см. ранее). Норэпинефрин действует в нескольких типах адренорецепторов: a1, a2, ?1-3. Большинство нейронов, содержащих норэпинефрин, в ЦНС расположены в голубоватом пятне в варолиевом мосту и среднем мозге. Их пути в других областях мозга показаны на рис. 8.27.
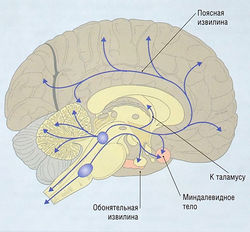
Рис. 8.27 Норэпинефрин в центральной нервной системе. Большая часть НЭ-содержащих нейронов в центральной нервной системе расположены в голубоватом пятне варолиевого моста и среднего мозга. Эти нейроны идут через связку средней части переднего мозга в лимбической системе к коре, таламусу и гипоталамусу. Вторая группа НЭ-содержащих нейронов в вентральной покрышечной области имеет отростки к гипоталамусу и миндалевидному телу.
5-гидрокситриптамин, депрессия и тревожное состояние
Норэпинефрин и аффективные и тревожные состояния
В нервной системе человека были идентифицированы пять типов дофаминовых рецепторов (D1-D5)
D1 D5-рецепторы стимулируют образование цАМФ, активизируя стимулирующий G-белок, в то время как D2-, D3- и 04-рецепторы ингибируют образование цАМФ, активизируя ингибирующий G-белок. D2-рецепторы более распространены, чем D3- и D4-рецепторы. D3-рецепторы преимущественно расположены в хвостатом ядре (одно из септальных ядер в лимбической системе), а D4-рецепторы сконцентрированы в медиальной лобной коре.
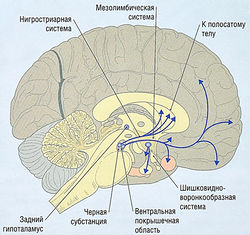
Рис. 8.28 Дофамин в центральной нервной системе. Дофаминергические тракты — нигростриарный, мезолимбический/мезокортикальный и шишковидно-воронкообразный.
Варианты дофаминергических трактов представлены на рис. 8.28:
Дофамин синтезируется как часть обычного пути для катехоламинов и метаболизируется двумя ферментами: МАОв, которая является интранейрональным ферментом, и катехол-O-метилтрансферазой, которая является экстранейрональным ферментом. Первичный метаболит дофамина — гомованиловая кислота.
Считается, что D2-рецепторы — наиболее важные дофаминовые рецепторы, которые вовлечены в развитие психоза, т.к. эффективность антипсихотических препаратов коррелирует с их сродством к D2-рецепторам. Однако появление нетипичных нейролептиков с равной эффективностью, но с относительно низким сродством к D2-рецептору повышает вероятность, что и другие подтипы дофаминовых рецепторов могут играть важную роль в этиологии и лечении психоза.
Постоянная блокада дофаминовых рецепторов ведет к их дезрегулированию, которое может вносить вклад в нарушения движения, обнаруживаемые при длительной нейролептической терапии.
Очевидно, что мезолимбический и мезокортикальный тракты играют важную роль в регулировании поведения, управляемого положительным подкреплением, и эти находки могут вести к созданию новых лекарственных средств для терапии физической зависимости.
Дофамин, болезнь Паркинсона и психоз
Известно около 300 пептидных нейромедиаторов в ЦНС
Пептиды имеют менее 100 аминокислот. Наиболее хорошо изученные нейропептиды перечислены в табл. 8.10. Часто эти пептиды синтезируются как часть больших молекул, названных предпрогормонами. Они расщепляются в нейрональной цитоплазме на прогормоны, которые затем собираются в пузырьки. В пределах пузырьков прогормоны далее расщепляются на нейроактивные пептиды. Большинство пептидных нейромедиаторов сосуществует с другими нейромедиаторами.
Считают, что опиоиды регулируют стресс, боль и настроение
Три эндогенные опиоидные группы, т.е. эндорфины, энкефалины и динорфины (см. табл. 8.10), синтезируются из больших молекул-предшественников. Эндорфин обнаружен в норадренергических и серотонинергических нейронах. Опиоиды действуют на три типа рецепторов:
Нейрогипофизарные нейроактивные пептиды вазопрессин и окситоцин могут быть вовлечены в регуляцию настроения
Два нейрогипофизарных гормона — вазопрессин и окситоцин — синтезируются в гипоталамусе и выделяются (секретируются) в заднюю долю гипофиза. Существуют три рецептора к вазопрессину, и его эффекты связаны с изменениями в фосфолипидах мембран или увеличением содержания цАМФ.
Тахикинины включают субстанцию Р и нейрокинин
Субстанция Р — первичный нейромедиатор в большинстве первичных афферентных чувствительных нейронов, присутствующий в нигростриарном тракте. Субстанция Р связана с АХ и 5-НТ и вовлечена в развитие хореи Хантингтона, болезни Альцгеймера и эмоциональных нарушений.
Холецистокинин может играть определенную роль в развитии шизофрении, панических расстройств, пищевых нарушений и некоторых расстройств движения
Холецистокинин (ХЦК) сосуществует в нейронах с дофамином и ГАМК. Он действует на два подтипа рецептора: ХЦКА и ХЦКВ. Подтип ХЦКА активирует эффект мембранных фосфолипидов. Механизмы сигнальной трансдукции ХЦКв-рецептора пока не выяснены.
Нейротензин может играть определенную роль в развитии шизофрении
Нейротензин сосуществует в нейронах с НЭ и дофамином. Нейротензин действует на G-белок, связанный с высокоаффинными рецепторами, находящимися в областях, богатых дофамином, и энтеро-обонятельной зоне коры, вовлеченных в развитие шизофрении.
Читайте также [ править ] НейромедиаторыЦель. Изучение влияния дилепта на регуляцию синтеза дофамина в условиях модели прерывания импульсной активности нейронов, вызванного введением гамма-бутиролактона. Методы. Содержание дофамина и его метаболитов в ткани мозга крыс определялось путем высокоэффективной жидкостной хроматографии с электрохимической детекцией. Результаты. Показано, что дилепт повышает уровень дофамина и его метаболитов гомованилиновой и диоксифенилуксусной кислот, а также ускоряет метаболизм дофамина в прилежащем ядре. Установлено также, что в условиях блокады импульсной активности дофаминергических нейронов дилепт повышает скорость синтеза дофамина. Выводы. Выявленный спектр фармакологической активности дилепта в отношении дофаминергической сис темы сходен с нейрохимическими эффектами, характерными для атипичных нейролептиков и эндогенного антипсихотика нейротензина .
Выпуск: 3 том 92 / 2011
Похожие темы научных работ по. автор научной работы — ШУБЕНИНА ЕВГЕНИЯ ВЯЧЕСЛАВОВНА, ОСТРОВСКАЯ РИТА УШЕРОВНА, КУДРИН ВЛАДИМИР СЕРГЕЕВИЧ, КЛОДТ ПЕТР МИХАЙЛОВИЧ, НАРКЕВИЧ ВИКТОР БОРИСОВИЧ, КУЗНЕЦОВА ЕЛЕНА АЛЕКСАНДРОВНА, ГУДАШЕВА ТАТЬЯНА АЛЕКСАНДРОВНА,Асимметрия в уровнях моноаминов в головном мозге мышей линии Balb/с, выращенныхв условиях социальной изоляции
2012 / Карпова Инесса Владимировна, Михеев Владимир Владимирович, Бычков Евгений Рудольфович, Лебедев Андрей Андреевич, Шабанов Петр ДмитриевичНейрофармакологический анализ эффектов оланзапина и клозапина на поведенческие характеристики и содержание нейромедиаторов в структурах мозга крыс
2004 / Гулиева С. Т. Ганиев М. М. Андреева Ю. А. Клодт П. М. Кудрин В. С. Наркевич В. Б. Раевский К. С.Структурные изменения в мезокортиколимбической дофаминергической системе мозга при длительной алкоголизации крыс
2008 / Дробленков А. В. Лебедев А. А. Шабанов П. Д.Роль дофамина в деятельности мозга (обзор литературы)
2010 / Циркин В. И. Багаев В. И. Бейн Б. Н.Действие ноотропного препарата дигам на обмен нейромедиаторов в мозге крыс при дисфункции дофаминергической системы
2008 / Доведова Елизавета Леонтьевна, Герштейн Лидия Михайловна, Ещенко Наталья Дмитриевна
УДК 616.895.8: 615.214.2: 612.085.1
ЭКСПЕРИМЕНТАЛЬНОЕ ИЗУЧЕНИЕ ВЛИЯНИЯ ДИПЕПТИДНОГО АНАЛОГА НЕЙРОТЕНЗИНА ДИЛЕПТА НА ДОФАМИНЕРГИЧЕСКИЕ СИСТЕМЫ ПРИЛЕЖАЩЕГО ЯДРА ГОЛОВНОГО МОЗГА
Евгения Вячеславовна Шубенина, Рита Ушеровна Островская*, Владимир Сергеевич Кудрин,
Петр Михайлович Клодт, Виктор Борисович Наркевич, Елена Александровна Кузнецова,
Татьяна Александровна Гудашева
НИИ фармакологии им. В.В. Закусова РАМН, г. Москва
Цель. Изучение влияния дилепта на регуляцию синтеза дофамина в условиях модели прерывания импульсной активности нейронов, вызванного введением гамма-бутиролактона.
Методы. Содержание дофамина и его метаболитов в ткани мозга крыс определялось путем высокоэффективной жидкостной хроматографии с электрохимической детекцией.
Результаты. Показано, что дилепт повышает уровень дофамина и его метаболитов гомованилиновой и диок-сифенилуксусной кислот, а также ускоряет метаболизм дофамина в прилежащем ядре. Установлено также, что в условиях блокады импульсной активности дофаминергических нейронов дилепт повышает скорость синтеза дофамина.
Выводы. Выявленный спектр фармакологической активности дилепта в отношении дофаминергической системы сходен с нейрохимическими эффектами, характерными для атипичных нейролептиков и эндогенного антипсихотика нейротензина.
Ключевые слова: дилепт, нейротензин, дофамин, прилежащее ядро, гамма-бутиролактоновая модель.
EXPERIMENTAL STUDY OF A DIPEPTIDE ANALOG OF NEUROTENSIN DILEPT ON THE DOPAMINERGIC SYSTEMS OF THE BRAIN NUCLEUS ACCUMBENS E.V. Shubenina, R.U. Ostrovskaya, V.S. Kudrin, P.M. Klodt, V.B. Narkevich, E.A. Kuznetsova, T.A. Gudasheva. Research Scientific Institute of Pharmacology in the name of V.V. Zakusov of the Russian Academy of Medical Sciences, Moscow city. Aim. To study the effects of Dilept on the regulation of synthesis of dopamine in a model of interruption of the impulse activity of neurons caused by the introduction of gammabutyrolaktone. Methods. The content of dopamine and its metabolites in rat brain tissue was determined by high performance liquid chromatography with electrochemical detection. Results: Show was the fact that Dilept increases the level of dopamine and its metabolites homovanillic and dioxyphenylacetic acids, and also accelerates the metabolism of dopamine in the nucleus accumbens. It was also established that Dilept increases the rate of synthesis of dopamine in the conditions of blockade of impulse activity of dopaminergic neurons. Conclusions: The established spectrum of pharmacological activity of Dilept on the dopaminergic system is similar to such of the atypical antipsychotics and to the endogenous antipsychotic neurotensin. Key words: dilept, neurotensin, dopamine, nucleus accumbens, the gammabutyrolaktone model.
Согласно дофаминергической гипотезе, ность повышать скорость оборота дофамина
позитивные и негативные симптомы ши- в прилежащем ядре и неостриатуме, пре-
зофрении связаны с гиперактивностью до- пятствовать развитию эффектов дофамино-
фаминергических нейронов в мезолимбиче- миметиков, потенцировать действие барби-
ской системе с одновременным снижением туратов, вызывать гипотермию, позволили
их функционирования в префронтальной сформулировать гипотезу о роли нейротен-
коре [10]. Большое значение в нарушениях зина в качестве эндогенного нейролептика
активности мезолимбической системы от- [7]. Достаточно сложная структура нейро-
водится изменению дофаминергической тензина, обусловливающая его низкую биоиннервации прилежащего ядра, актива- доступность для мозга, является причиной
ция которого ведет к подавлению тонуса того, что эффекты нейротензина воспроиз-
соответствующей системы коры [12]. Ха- водятся только при введении этого тридека-
рактерным свойством классических и ати- пептида в желудочки мозга или при подве-
пичных антипсихотических препаратов дении его к определенным структурам [4].
является способность ускорять метаболизм Это исключает возможность клинического
дофамина в мезолимбической системе. применения нейротензина в качестве ан-
Этот эффект принято рассматривать как типсихотического средства. Оригинальный
компенсаторный ответ, развивающийся подход к поиску высокоактивных пептид-
по механизму обратной связи в результате ных препаратов, развиваемый в НИИ фар-
блокады пресинаптических дофаминовых макологии имени В.В. Закусова РАМН, со-
рецепторов Д2 подтипа [3]. Исследования стоит в создании замещенных дипептидов,
показали, что подобный эффект наблюда- имитирующих структуру активного центра
ется и у нейропептида нейротензина (НТ). эндогенного пептида и непептидного ле-
Такие эффекты нейротензина, как способ- карственного препарата с соответствующей
------------------------- активностью. За основу моделирования ак-
* Автор для переписки: niipharm@mail.ru тивных НТ-подобных нейролептиков был
взят дипептид пролил-тирозин в связи с тем, что эта последовательность соответствует центральному фрагменту изгиба основного метаболита нейротензина, НТ8-13 и имеет топологическое сходство с атипичным нейролептиком сульпиридом [6]. Из серии №ацил-пролил-тирозинов для более подробного изучения был отобран №капроил-Ь-пролил-Ь-тирозина метиловый эфир (ди-лепт), продемонстрировавший активность по стандартным показателям нейролептического действия. На основе полученных результатов было высказано предположение, что дилепт в условиях системного введения воспроизводит поведенческие эффекты, характерные для нейротензина, вводимого в желудочки мозга. Для выяснения механизмов данного явления представляется перспективным нейрохимический анализ активности дилепта. Наряду с изучением изменений исходной активности дофаминовой системы в ткани прилежащего ядра целесообразно применение функциональной модели прерывания импульсной активности нейронов. Использование гамма-бути-ролактона (ГБЛ) в этой модели позволяет блокировать импульсную активность дофа-минергических нейронов, относящихся к проекции черной субстанции в прилежащее ядро, резко усиливать синтез дофамина, подавлять его высвобождение в межсинапти-ческое пространство, за счет которого повышается способность пресинаптических Д2 рецепторов связываться с предполагаемыми лигандами. Дополнительное введение ингибитора декарбоксилазы ароматических аминокислот 3-оксибензилгидразина (3-ОБГ) в ГБЛ модели позволяет судить о влиянии исследуемого вещества на скорость синтеза предшественника дофамина — ди-оксифенилаланина (ДОФА), определяемой активностью тирозингидроксилазы. Активность этого фермента находится в обратной зависимости от сигналов, исходящих от пресинаптических дофаминовых рецепторов [11].
Цель настоящей работы заключалась в изучении влияния дилепта на исходный уровень дофамина и его метаболитов в ткани прилежащего ядра крыс Вистар, а также в оценке его действия на синтез дофамина в условиях ГБЛ модели.
Работа была выполнена на 48 крысах самцах линии Вистар массой 250 — 300 г. Были проведены две серии экспериментов. Задача первой серии заключалась в изучении влияния дилепта на содержание
катехоламинов и их метаболитов в ткани прилежащего ядра крыс Вистар. Крыс подразделили на четыре группы по 8 животных в каждой. В первой и второй группах вводили дилепт (0,8 мг/кг) внутрибрюшинно (в/б), в третьей и четвертой — физиологический раствор (в/б) в эквивалентном объеме. Декапитация животных первой и третьей группы осуществлялась через 30 минут после введения препарата или физиологического раствора, а во второй и четвертой — через 60 минут. Прилежащее ядро извлекали на льду, замораживали в жидком азоте и гомогенизировали в 500 мкл среды выделения, содержащей 0,1н НСЮ4 с 0,5 нмоль/мл диоксибензиламином в качестве внутреннего стандарта. После этого пробы центрифугировали при 10000 g в течение 15 минут. В дальнейшем супернатант использовали для определения содержания предшественника дофамина ДОФА методом высокоэффективной жидкостной хроматографии с электрохимической детекцией (ВЭЖХ-ЭД) [1].
Задачей второй серии исследований было изучение влияния дилепта на синтез дофамина в прилежащем ядре крыс линии Вистар в условиях ГБЛ модели. 16 крыс распределяли по двум группам поровну. В первой группе за 60 минут до декапитации животным вводили дилепт (0,8 мг/кг) внутрибрюшинно (в/б), а во второй — физиологический раствор (в/б) в эквивалентном объеме. После введения дилепта либо физиологического раствора животным обеих групп внутрибрюшинно вводили ГБЛ (750 мг/кг) и 3-ОБГ (100 мг/кг) соответственно за 35 и 20 минут до декапитации. Извлечение прилежащего ядра и последующие экспериментальные процедуры проводились в соответствии с протоколом первой части эксперимента.
Статистическую обработку полученных данных производили по программе Statistica 6.0. Достоверность различий оценивали с помощью ^критерия Стьюдента.
Через 30 минут после введения дилепта в дозе 0,8 мг/кг уровень дофамина (ДА) и его метаболитов — диоксифенилуксусной кислоты (ДОФУК) и гомованилиновой кислоты (ГВК) составил 118±3%, 126±5%,125±3% соответственно по сравнению с таковыми в группе крыс, которым вводился физиологический раствор (контроль). Комплексные показатели ГВК/ДА и ДОФУК/ДА, характеризующие скорость утилизации дофамина, составляли 143±8% и 135±7% соответствен-
но. Через 60 минут после введения дилепта соотношение ДОФУК/дофамин возрастало до 127±4% по сравнению с контролем.
Как следует из представленных данных, дилепт ускоряет оборот дофамина в прилежащем ядре. Достоверные изменения уровня дофамина и его метаболитов выявляются в основном через 30 минут после введения препарата, а через 60 минут устойчивые изменения касаются только соотношения ДОФУК/ДА. Известно, что типичные нейролептики, например галоперидол, увеличивают оборот дофамина в прилежащем ядре более чем на 50% [2, 9], тогда как атипичные нейролептики, такие как суль-пирид, повышают этот показатель только на 15 — 20% [2]. Эффект, характерный для атипичных нейролептиков, описан и для эндогенного нейролептика нейротензина [9]. Таким образом, можно сделать вывод, что дилепт увеличивает скорость оборота дофамина в степени, характерной для эффектов атипичных антипсихотиков и ней-ротензина.
Повышение уровня ДОФА в условиях ГБЛ модели до детектируемого позволило оценить изменение синтеза дофамина после введения дилепта. За 100% был принят уровень ДОФА, измеренный после введения физиологического раствора на фоне 3-ОБГ. Как показали наши исследования, введение ГБЛ в сочетании с ингибитором ароматических кислот повышало содержание ДОФА в прилежащем ядре, что полностью соответствует данным литературы [11]. Нами показано увеличение содержания ДОФА до 324±20%. Введение дилепта приводило к еще большему достоверному возрастанию этого показателя до 405±41%, что свидетельствовало об усилении синтеза дофамина.
Компенсаторное усиление синтеза дофамина в дофаминергическом нейроне по принципу обратной связи может быть ответом на блокаду Д2 рецепторов дофамина [13]. Такое изменение ведет, в свою очередь, к активации метаболизма дофамина. ГБЛ модель создаёт условия для изучения способности веществ воздействовать на накопление ДОФА, исключая их влияние на высвобождение и метаболизм дофамина. Показано, что в условиях ГБЛ модели агонисты дофамина снижают уровень ДОФА, а антагонисты усиливают его накопление в прилежащем ядре [11]. Нейротензин в условиях ГБЛ модели также усиливает синтез дофамина [9]. Более позднее изучение это-410
го нейропептида показало, что он обладает способностью блокировать рецепторы дофамина Д2 типа [3] и тем самым усиливает активность тирозингидроксилазы [8]. Исследования, проведенные A. Drumheller и A. Reches [5, 9], показали, что нейротензин повышает величину соотношения ДОФУК/ ДА и ГВК/ДА в прилежащем ядре, следовательно, ускоряет утилизацию дофамина в мезолимбической системе. Исходя из этого можно предположить, что одним из механизмов нейролептикоподобного действия дилепта является его способность усиливать синтез и ускорять метаболизм дофамина. Нельзя исключить, что способность дилеп-та увеличивать синтез дофамина связана с его антагонистическим влиянием на пре-синаптические Д2 рецепторы, следствием которого является повышение активности тирозингидроксилазы, однако подобные предположения требуют дальнейшего изучения.
Полученные данные о способности ди-лепта повышать содержание дофамина и его метаболитов в ткани прилежащего ядра и увеличивать синтез дофамина в условиях ГБЛ модели свидетельствуют о сходстве нейрохимических механизмов действия этого дипептида и атипичных антипсихотиков, а также нейротензина.
1. Кудрин В.С. Мирошниченко И.И. Раевский К.С. Различия в механизмах ауторецепторной регуляции биосинтеза и высвобождения дофамина в подкорковых структурах мозга крыс // Нейрохимия. — 1988. — Т.7. — №.1. — С. 3 — 8.
2. Мирошниченко И.И. Кудрин В.С. Раевский К.С. Влияние карбидина, сульпирида и галоперидола на содержание моноаминов и их метаболитов в структурах головного мозга крыс // Фармакол. и токсикол. — 1988. — Т. 51.- № 2. — С. 26 — 29.
3. Binder E.B. Kinkead B, Owens M.J. Nemeroff C.B. Neurotensin and dopamine interactions // Pharmacol. Rev. —
2001. — Vol. 53. — № 4. — P. 453 — 486.
4. Boules M, Fredrickson P. Richelson E. Bioactive analogs of neurotensin: focus on CNS effects // Peptides. — 2006. — Vol. 27. — №10. — P. 2523 — 2533.
5. Drumheller A.D. Gagnu M.A. St-Pierre S. Jolicoe-ur F.B. Effects of neurotensin on regional brain concentrations of dopamine, serotonin and their main metabolites // Neuropeptides. — 1990. — Vol. 15. — № 3 — P. 169 — 178.
6. Gudasheva T.A. Voronina T.A. Ostrovskaya R.U. et al. Design of N-acylprolyltyrosine «tripeptoid» analogues of neurotensin as potential atypical antipsychotic agents // J. Med. Chem. — 1998. — Vol. 41. — № 3. — P. 284 — 290.
7. Kinkead B, Nemeroff C.B. Neurotensin: an
endogenous antipsychotic? // Curr. Opin. Pharmacol. —
2002. — Vol. 2. — №1 — P. 99 — 103.
8. Najimi M, Robert J.J. Mallet J. et al. Neurotensin induces tyrosine hydroxylase gene activation through nitric
oxide and protein kinase C signaling pathways // Mol. Pharmacol. - 2002. - Vol. 62. - № 3. - P. 647 - 53.
9. Reches A, Burke R.E. Jiang D, et al. Neurotensin interacts with dopaminergic neurons in rat brain // S. Peptides. - 1983. - Vol. 4. - № 1. - P. 43 - 48.
10. Tzschentke TM. Pharmacology and behavioral pharmacology of the mesocortical dopamine system // Prog. Neurobiol. - 2001. - Vol. 63. - № 3. - P. 241 - 320.
11. Walters J.R. Roth R.H. Dopaminergic neurons -alteration in the sensitivity of tyrosine hydroxylase to
inhibition by endovenous dopamine after cessation of impulse flow // Biochem Pharmacol. - 1976. - Vol. 25. -№ 6. - P. 649 - 654.
12. Weinberger D.R Implications of normal brain development for the pathogenesis of schizophreni. // Arch. Gen. Psychiatry. - 1987. - Vol. 44. - №7. - P. 660 - 669.
13. Yoshida S.T. Baella S.A. Stuebner N.M. et al. Effects of a partial D2-like receptor agonist on striatal dopamine autoreceptor functioning in preweanling rats // Brain Res. -2006. - Vol. 10 73 - 1074. - P. 269 - 275.
УДК 616.153.915: 615.322: 615.272.4: 612.084
ИССЛЕДОВАНИЕ ГИПОЛИПИДЕМИЧЕСКОЙ АКТИВНОСТИ МАСЛЯНОГО ЭКСТРАКТА ИЗ КОРНЯ ЛОПУХА
Анес Габбасович Беккужин, Айман Кажигалеевна Бердгалеева * Западно-Казахстанский государственный медицинский университет, г. Актобе, Республика Казахстан
Цель. Изучение гиполипидемической активности фитопрепарата масляного экстракта из корня лопуха при экспериментальной гиперхолестеринемии у крыс.
Методы. Эксперименты проводились на 30 белых крысах, у которых воспроизводили модель холестериновой гиперлипидемии. Масляные экстракты из корня лопуха вводили внутрижелудочно в дозе 2,5 мл/кг в течение 2 месяцев. После выведения животных из эксперимента в гомогенатах печени с помощью тонкослойной хроматографии определялось относительное содержание фракций фосфолипидов.
Результаты. Введение масляного экстракта из корня лопуха на фоне холестеринового атеросклероза снижало уровень лизофосфатидилхолина по сравнению с моделью патологии в 3 раза. Фракции фосфатидилхолина, суммы фосфитилсерина и сфингомиелина, фосфатидилинозина, сульфатидов и фосфатидной кислоты у животных, получавших препарат, достоверно повышались по сравнению с моделью холестеринового атеросклероза соответственно на 62%, 11%, 30%, 38% и78%.
Выводы. Введение масляного экстракта из корня лопуха животным, получавшим холестерин, способствует восстановлению спектра фосфолипидов печени.
Ключевые слова: фитоэкстракты, холестериновый атеросклероз, фосфолипиды печени.
THE STUDY OF THE HYPOLIPIDEMIC PROPERTIES OF THE OIL EXTRACT FROM THE BURDOCK ROOT
A.G. Bekkuzhin, A.K. Berdgaleeva, West-Kazakhstan State Medical University, Aktobe city, Republic of Kazakhstan Aim. To study the hypolipidemic activity of the oil extract phytopreparation of the burdock root in experimental hypercholesterolemia in rats. Methods. Experiments were performed on 30 white rats, in which the model of cholesterol hyperlipidemia was reproduce. Oil extracts of the burdock root were administered intragastrically at a dose of 2.5 ml/kg for 2 months. After removing the animals from the experiment the relative content of phospholipid fractions was determined in liver homogenates by using the thin layer chromatography technique. Results: The introduction of an oil extract of the burdock root for cholesterol atherosclerosis decreased the level of lysophosphatidylcholine in comparison with the pathological model by 3 times. Fractions of phosphatidylcholine, the content of phosphitylserin and sphingomyelin, phosphatidylinozin, sulfatides and phosphatidic acid in the animals receiving the drug, significantly increased compared with the model of cholesterol atherosclerosis, respectively. Conclusions: Administration of an oil extract of the burdock root to animals receiving cholesterol, helps to restore the spectrum of phospholipids of the liver. Key words: phytoextracts, cholesterol atherosclerosis, liver phospholipids.
Ишемическая болезнь сердца (ИБС), обусловленная атеросклерозом, остается основной причиной смертности во всем мире [1]. Доказано, что повышенный риск развития ИБС и других проявлений атеросклеротического поражения сосудов связан с увеличением содержания ХС в крови, особенно в виде ХС липопротеи-дов низкой плотности (ЛПНП). Лучшее лечение атеросклероза — его профилактика, что предусматривает соблюдение здорового образа жизни и соответствующей диеты. Современные методы лечения на-
* Автор для переписки: bak0803@mail.ru
правлены главным образом на снижение концентрации ЛПНП и ЛПОНП, а также на повышение концентрации антиатеро-генных ЛПВП [2]. Медикаментозная терапия атеросклероза сопровождается рядом побочных действий, а эффективна лишь в 10% случаев [3]. В последние годы возрос интерес к лекарственным препаратам растительного происхождения, что обусловлено прежде всего их низкой токсичностью и относительной безопасностью [4, 5]. Среди препаратов растительного происхождения приобретают актуальность жирорастворимые фитоэкстракты.
Нами был изучен разработанный на кафедре фармакологии ЗКГМУ им. М. Ос-